Location:Home >> Detail
Med One. 2016; 1(2): 2; https://doi.org/10.20900/mo.20160007

1 Department of Neurosurgery, Xiangya 3rd Hospital, Changsha, Hunan 410013, P.R. China
2 Department of Neurosurgery, Xianyang Central Hospital, Xianyang, Shaanxi 712000, P.R. China
3 American Informatics Consultant LLC, Rockville, MD 20852, USA
*Corresponding Author: Lydia C Manor.
Background: Glioblastoma Multiforme (GBM) is a common malignant brain tumor. It is characterized by rapid growth and high tumor heterogeneity. The molecular mechanisms driving GBM tumorigenesis and progression are not yet fully elucidated.
Methods: miR-124, and miR-137, expression in 43 high-grade GBM tumor tissues was measured by real-time PCR and compared with expression in 24 normal brain tissues. The effects of miR-124, and miR-137, overexpression on glioma U251 cell proliferation and differentiation were analyzed. Tuj1 and GFAP expression was detected using immunofluorescence staining. Cell proliferation was detected with flow cytometry.
Results: miR-124, and miR-137, expression was significantly decreased in GBM tumor tissues compared to normal brain tissues (p < 0.01). Immunofluorescence showed, after miR-124 and miR-137 transfection, normal synapse growth structure in GBM, and that neuronal differentiation factor expression significantly increased, including Tuj1 and GFAP. Flow cytometry analysis showed that GBM cell cycle extension and differentiation was repressed.
Conclusion: miR-124, and miR-137, function as tumor suppressors to inhibit cell proliferation and differentiation and in GBM, their expression was significantly decreased.
Glioblastoma Multiforme (GBM) is a common malignant tumor, accounting for 80 % of adult, unknown etiology, primary malignant brain tumors [1]. GBM is divided, using its pathological and clinical features, into astrocytoma, oligodendroglioma, oligoastrocytomas, and ependymoma [2]. GBM is characterized by rapid growth, high tumor heterogeneity, and diffuse infiltrative growth. These make treatment difficult and results in high recurrence, and poor survival, rates [3]. Current GBM treatment therapy is based on surgery combined with radiotherapy, chemotherapy, immunotherapy, and molecular targeting. This therapy regimen can improve GBM patient prognosis, but fails to provide a radical cure of GBM [4]. GBM cell proliferation, and differentiation, regulatory systems are of great significance to prolonging patient survival and improving prognosis.
A microRNA (miRNA) is a conserved, small, non-coding RNA molecule ( ~ 18-25 nucleotides long), which functions in the post-transcriptional regulation of gene expression by binding to the 3'-UTR of a target mRNA and causing subsequent cleavage, destabilization, and less efficient mRNA translation [5, 6]. As of 2016, over 1,000 miRNAs have been found in humans, and is proposed to target about 60 % of all genes [7, 8]. Neural differentiation is characterized with the expression of specific proteins, including Tuj1 and GFAP, which are essential for cytoskeleton and associated with cellular morphology [9, 10]. miR-124, and miR-137, overexpression results in changes in cell proliferation and differentiation [11]. Human β-Tubulin 3 is a structural protein (450 amino acid) expressed in neurons and is thought to be a neuron specific class III tubulin (Tuj1). It contributes to microtubule stability in neuronal cell bodies and axons [12]. The glial fibrillary acidic protein (GFAP) is an intermediate filament protein that expresses in numerous cell types of the central nervous system, including astrocytes and ependymal cells [13]. GFAP is involved in many important CNS processes, including astrocyte-neuron interactions, cell-cell communication, blood brain barrier function maintenance, and CNS injury repair [14, 15]. This study investigated miR-124, and miR-137, expression in GBM and their effects on proliferation and differentiation, and then examined Tuj1 and GFAP expression in GBM cells.
A total of 43 high-grade GBM tumor tissue samples were collected, between December 2012 and December 2014, from patients before chemotherapy, radiotherapy, or other treatments. GBM was diagnosed using head MRI and pathological examinations. Of the 43 cases, 23 were male and 20 were female with an average age of 39.1 ± 6.5. There were 24 relatively normal brain tissues collected around the trauma of cerebral internal injuries as controls. Of the 24 controls, 13 were male and 11 were female with an average age of 41.3 ± 8.2. Tumor, and control brain, tissues were obtained during surgery and immediately placed in liquid nitrogen. This study was approved by ethics committee. Signed consent forms were collected from all study subjects.
2 Cell cultureThe U251 glioma cell line was provided by the Chinese Academy Sciences Cell Bank (China) and cultured, at 37°C, 5 % CO2, in an RPMI 1640 medium containing 10 % fetal bovine serum (FCS), 4 × nonessential amino acids, 3 mM L-Glutamine, Penicillin (100 U/ml), and, Streptomycin (100 mg/ml).
2.3 Real-time RT-PCRmiR-124, and miR-137, expression levels in GBM tumor tissues and normal brain tissues were detected by using a Real-time RT-PCR. First, primers were designed based on an miR-124 and miR-137 sequence (GeneBank accession numbers: NR_0296 68, and NR_029679, respectively) (Table 1). RNA was extracted from tissues with an RNAprep Pure Tissue Kit. Real-time RT-PCR was performed by a Real-time PCR Amplifier (Bio-Rad) using an mirVanat qRT-PCR miRNA kit (Ambion) under the following reaction conditions: 95°C, for 3 min; 95°C for 15 s; and 60°C at 30 s for 40 cycles. U6 was used as a reference gene. Results were analyzed using 2-ΔΔCt [16].
 Table 1. Synthesis of nucleic acid sequence 2 for cell transfection
Table 1. Synthesis of nucleic acid sequence 2 for cell transfection
Matured miR-124, and miR-137, sequences were used to establish an miR-124, and miR-137, overexpression system. The corresponding sequences with the same oligonucleotide sequences compositions were used as negative controls. miR-124, and miR-137, sequences were synthesized by the Nanjing Shengxing Company. After cloning the synthesized miRNA sequences into the pMIR-REPORT expression system (Ambion), the plasmid DNA with correct insertion WAS transfected into U251 glioma cells using INTERFERinTM Polyplus transfection reagents. THE U251 GBM cell line was purchased from the Cell Bank of the Chinese Academy of Science. Cells were cultured in an RPMI-1640 medium (HyClone) at 37°C and 5 % CO2 for 24 hrs after transfection. Cells were then subjected to further analysis.
2.5 ImunofluorescenceImmunofluorescence was used to detect Tuji and GFAP expression levels in miRNA-expression-vector-transfected U251 cells. Logarithmic growth phase transfected cells were fixed with paraffin onto slides. After blocking, cells were incubated with mouse anti-human an Tuji and GFAP (1:2000) antibody for 2 hrs at 37°C, followed by incubation with a fluorescence-marked goat anti-mouse secondary antibody (1:1000) for 1 hr at 37°C. After washing with 1 × PBS and attaching cover slips to the slices, fluorescent staining was observed under a fluorescence microscope.
2.6 Flow cytometry assaymiR-124 and miR-137-transfected U251 glioma cells were grown to the logarithmic phase (24 hrs) and fixed with 90 % ethanol, overnight, at 4°C. After removing the ethanol, cells were incubated with RNase for 30 min at 37°C and cell nuclei were stained with PI. The stained cells were then subjected to a flow cytometry assay (Becton Dickinson) with an excitation wavelength of 488 nm and an emission wavelength of 630 nm. The FL-2 area, and DNA histograms, were analyzed by Modifit software. All experiments were repeated 3 times.
2.7 Statistical AnalysisResults were analyzed using SPSS 20.0 and presented with a mean ± standard deviation. T test was used for comparative analysis. p < 0.05 was deemed as statistically significant.
miR-124, and miR-137, expressions in GBM was examined with RT-PCR (Fig. 1). miR-124, and miR-137, expression in GBM tissues significantly decreased compared to normal brain tissue (p < 0.01). miR-124, and miR-137, expression in normal brain tissue was 12 and 15 times higher, respectively, than that in GBM tumor tissues.
 Fig. 1 Relative expression of miR-124 and miR-137.
Fig. 1 Relative expression of miR-124 and miR-137.
miR-124 and miR-137 expression in normal brain tissues and GBM tumor tissues was measured by real-time PCR. *p < 0.01 between normal brain tissues and GBM tumor tissues.
A pMIR-REPORT expression system was used to detect miR-124, and miR-137, overexpression in U251 glioma cells. Total RNA was extracted. miR-124, and miR-137, expression was measured using real-time PCR. miR-124 and miR-137, relative expression is shown in Fig. 2. Cell transfection significantly increased miR-124, and miR-137, expression in U251 glioma cells compared to control cells (p < 0.05).
 Fig. 2 Overexpression of miR-124 and miR-137 in U251 cells.
Fig. 2 Overexpression of miR-124 and miR-137 in U251 cells.
Glioma U251 cells were trransfected with miR-124 and miR-137 expression vector, negative control vector (control), and without vector (sham). 24 hrs after the transfection, miR-124 and miR-137 expression in transfected cells were measured by real-time PCR. p < 0.05 between miR-124 or miR-137 trransfected cells and control cells.
miR-124, and miR-137, transfected U251 cells were harvested every other day. Tuj1, and GFAP, expression was detected by immunofluorescence staining (Fig. 3). Tuj1 was shown with GFP (a green fluorescent marker), GFAP with rhodamine (a red fluorescent marker). Tuj1 and GFAP cellular expression levels represent the degree of cell differentiation as specific proteins of the neuron system. With cell culture progression, Tuj1, and GFAP, expression gradually increased after miR-124, and miR-137, transfection. Phase contrast microscopy showed neuron-specific synapse structures in U251 cells after miR-124, and miR-137, overexpression, suggesting that the U251 cells differentiated into normal neurons.
 Fig. 3 Immunofluorescence of Tuj1 and GFAP expression.
Fig. 3 Immunofluorescence of Tuj1 and GFAP expression.
U251 cells were transfected with miR-124 or miR-137 expression vectors. Tuj1 (green) and GFAP (red) expression was detected by immunofluorescence staining (400×). A time-dependent increase in fluorescence intensity (expression level) was observed.
The cell growth cycle was measured using the flow cytometry of miR-124, and miR-137, which had transfected, blank-controlled, and negative-controlled U251 cells. FL-2 areas were analyzed using Modifit software (Fig. 4A). Cell distribution in different cell cycle stages was analyzed using SPSS 20.0 software (Fig. 4B, 4C). No differences were observed between: blank, and negative, G0/G1 phase control cells; S phase; or G2/M phase (p > 0.05). miR-124, and miR-137, transfection significantly increased the number of U251 cells in the G0 / G1 phase and decreased the number of U251 cells in the S, and G2/M, phases (p < 0.05), suggesting that cell cycle was prolonged and mitosis attenuated.
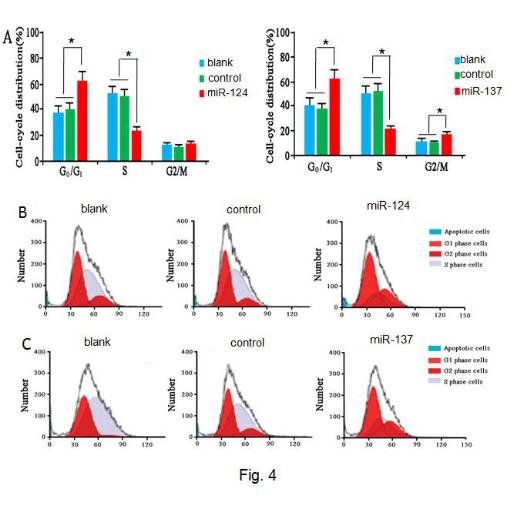 Fig. 4 Flow cytometry assay of cell cycle of miR-124 and miR-137 overexpressed U251 cells.
Fig. 4 Flow cytometry assay of cell cycle of miR-124 and miR-137 overexpressed U251 cells.
(A) Cell cycle distribution. *p < 0.05 between miR-124 or miR-137 trransfected cells and control cells. (B) Representative photogram of the number of cells in each cell cycle in U251 cells transfected with miR-124 vector, negative vector, and without vector. (C) Representative photogram of cell cycle distribution in U251 cells transfected with miR-137 vector, negative vector, and without vector.
This study first found significantly decreased miR-124, and miR-137, expression in high-grade GBM tumor tissues. Further investigation of glioma U251 cells, using an established overexpression model, confirmed miR124, and miR-137, effects in cell proliferation. Immunofluorescence staining revealed that miR124 and miR-137 overexpression stimulated neuronal differentiation. miR-124, and miR-137, abnormalities have been previously reported in GBM. But their biological functions have not been fully elucidated. Santos et al. demonstrated that low miR-124, and miR-137, expression is associated with GBM cell proliferation by maintaining activation of growth factors, such as EGF and FGF [17]. miR-124 and miR-137 decreased expression in GBM is proposed to result from irreversible modification of transcription regulation. For example, Chakrabarti et al. showed that miR-124 expression level was elevated in GBM after a DNA demethylation (hypomethylation) modifier, 5-Aza-2-deoxycytidine treatment [18]. This study provided no evidence as to the cause of low miR-124, and miR-137, expression in GBM tumor tissues, it demonstrated that miR-124, and miR-137, expression was significantly lowered in high grade GBM tumor tissues. Cell culture experiments with artificial expressions of these two miRNAs significantly inhibited glioma U251 cell proliferation. This study suggests that both miR-124, and miR-137, are GBM tumor suppressors. The roles of miR-124 and miR-137 in neuronal differentiation have been previously observed. miR-124 overexpression can enhance embryonic stem cell differentiation into neurons [19]. miR-124 was found to regulate PTBP1 expression, which plays a role in inhibiting alternative splicing patterns of mRNA precursors in non-neuronal cells [20]. Research has also found that miR-124 promotes neuronal differentiation by inhibiting SCP1 expression [21]. miR-137 was reported to inhibit CDK6 expression and subsequently participate in cell cycle, and mitosis, regulation [22]. This study found that miR-124 and miR-137 overexpression promotes U251 glioma cells differentiation into normal neurons and is accompanied by Tuj1, and GFAP, expression elevation.
As a severe brain tumor, GBM threatens human health, making diagnosis of, and treatment for, GBM an issue of common concern in clinical scenarios [1]. The instant study proved that miR-124 and miR-137 expression significant decreased in GBM cells, thus providing evidence for clinical diagnosis. It was also found that miR-124, and miR-137, over-expression induced GBM differentiation. Given that differentiation therapy is currently of great interest to the tumor field, as it induces cancer cells to transform with normal morphology, gene expression and function [22], the findings of this study will contribute to an improvement in differentiation therapy.
The authors declare no conflict of interests.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
Wu H, Liu Z, Manor L C. Loss of miR-124 and miR-137 Expression in Glioblastoma Multiforme and Their Roles in Glioma Cell Proliferation and Differentiation. Med One. 2016; 1(2): 2; https://doi.org/10.20900/mo.20160007

Copyright © 2020 Hapres Co., Ltd. Privacy Policy | Terms and Conditions



